Ronald Palacios Castrillo
RESUMEN
Dos sistemas de control de comportamiento están intercalados en la corteza motora del humano. Un sistema bien conocido consta de circuitos específicos de efectores para movimientos precisos y aislados de apéndices altamente especializados (dedos de manos y pies y lengua), el tipo de movimiento diestro necesario para hablar o manipular objetos.
Un segundo sistema integrador de resultados,la red de acción somatocognitiva (SCAN) , es más importante para controlar el organismo como un todo. El SCAN integra el control corporal (motor y autonómico) y la planificación de la acción, de acuerdo con la idea de que los aspectos del control ejecutivo de nivel superior pueden derivar de la coordinación del movimiento.
El SCAN incluye regiones específicas de M1, área motora suplementaria (SMA), tálamo (VIM y CM), putamen posterior y el cerebelo postural, y está funcionalmente conectado a regiones corteza cingulada anterior dorsal (dACC) vinculadas al libre albedrío, regiones parietales que representan intenciones de movimiento y regiones insulares para procesar señales somatosensorial, dolor y señales viscerales interoceptivas.
=> Recibir por Whatsapp las noticias destacadas
La aparente expansión relativa de las regiones SCAN en los seres humanos podría sugerir un papel en acciones complejas específicas de los seres humanos, como la coordinación de la respiración para el habla y la integración del movimiento de manos, cuerpo y ojos para el uso de herramientas.
Un factor común en esta amplia gama de procesos es que deben integrarse si un organismo quiere lograr sus objetivos a través del movimiento mientras evita lesiones y mantiene la alostasis fisiológica. El SCAN proporciona un sustrato para esta integración, permitiendo cambios posturales, respiratorios, cardiovasculares y de excitación anticipatorios previos a la acción (como tensión en los hombros, aumento del ritmo cardíaco o «mariposas en el estómago»).
El hallazgo de que la acción y el control del cuerpo se fusionan en un circuito común podría ayudar a explicar por qué los estados de la mente y el cuerpo interactúan con tanta frecuencia.
Esto demuestra el brillante trabajo de Gordon y colaboradores resumido abajo.
TEXTO
Una red de acción somatocognitiva se alterna con regiones efectoras en la corteza motora
Gordon, E.M., Chauvin, R.J., Van, A.N. et al. A somato-cognitive action network alternates with effector regions in motor cortex. Nature (2023). https://doi.org/10.1038/s41586-023-05964-2
A partir de la década de 1930, Penfield y sus colegas mapearon M1 humano con estimulación cortical directa, provocando movimientos de aproximadamente la mitad de los sitios, principalmente del pie, la mano y la boca.
Aunque las representaciones de partes específicas del cuerpo se superpusieron sustancialmente, estos mapas dieron lugar a la vista de libros de texto de la organización M1 como un homúnculo continuo, de pies a cabeza. En los primates no humanos, se han descrito características organizativas inconsistentes con los homúnculos del motor.
Los estudios de conectividad estructural dividieron M1 en motor grueso anterior, M1 «antiguo» (pocas proyecciones directas a las neuronas motoras espinales) y posterior, motor fina, «nuevo» M1 (muchas proyecciones motoneuronas directas). Los estudios de estimulación de primates no humanos mostraron que el cuerpo estaba representado en M1 anterior y los efectores motores (cola, pie, mano y boca) en M1 posterior.
Dichos estudios también sugirieron que las extremidades están representadas en zonas funcionales concéntricas que avanzan desde los dígitos en el centro hasta los hombros de la periferia.
Además, las estimulaciones podrían provocar acciones cada vez más complejas y multiefectoras al pasar de la M1 posterior a la anterior. Durante el comportamiento natural, los movimientos voluntarios forman parte de acciones dirigidas a objetivos, iniciadas y controladas por regiones ejecutivas en la CON.
Actividad neural anterior Los movimientos voluntarios pueden detectarse primero en la zona cingulada rostral dentro de la corteza cingulada anterior dorsal (DACC), luego en el área motor pre suplementaria (pre-SMA) y el área motor suplementaria (SMA), seguido de M1.
Todas estas regiones se proyectan a la médula espinal, con M1 como principal transmisor de motores de mandos por el tracto corticoespinal.
Las copias motoras eferentes son recibidas por la corteza somatosensorial primaria (S1), el cerebelo y el striatum para la corrección en línea, el aprendizaje y la inhibición de los movimientos competidores. Las inyecciones de trazadores en primates no humanos demostraron proyecciones desde M1/CON anterior hasta órganos internos (como la médula suprarrenal) para la excitación simpática preparatoria en previsión de la acción.
Los errores posteriores al movimiento y las señales de dolor se transmiten principalmente a las regiones insulares y cinguladas de la CON, que actualizan los planes de acción futuros.
La conectividad funcional en estado de reposo (RSFC) fMRI mapea de forma no invasiva las redes funcionales del cerebro. Los estudios de mapeo funcional de precisión (PFM) se basan en grandes cantidades de datos multimodales (como RSFC y tareas) para mapear la organización del cerebro específico del individuo con el mayor detalle posible.
Los primeros estudios de PFM identificaron regiones M1 separadas de pies, manos y boca con sus respectivos objetivos cerebelosos y estriados. Estos circuitos de motor de pie, mano y boca se caracterizaron por una fuerte conectividad dentro del circuito y especificidad del efector en la tarea FMRI, de acuerdo con la evidencia mieloarquitectónica para distintos campos corticales.
Sin embargo, estos circuitos estaban relativamente aislados y no incluían conexiones funcionales con redes de control como CON que podrían apoyar la integración del movimiento con los objetivos de comportamiento globales. Un estudio reciente mostró que la inmovilización de brazos dominante prolongada fortaleció la conectividad funcional entre M1 en desuso y el CON, lo que sugiere que el papel de CON puede extenderse más allá del control de la acción abstracta y de la coordinación del movimiento. Aquí utilizamos la última iteración de PFM con mayor resolución (2,4 mm) y mayores cantidades de fMRI (RSFC: 172–1813 min por participante; tarea: 353 min por participante) y datos de difusión, para mapear M1 y sus conexiones con el mayor detalle.
Los resultados se verificaron en datos promediados por grupo de los tres estudios de fMRI más grandes (Human Connconcome Project (HCP), Adolescent Brain Cognitive Development (ABCD), UK BioBank (UKB), total de n de aproximadamente 50.000. Además, colocamos nuestros hallazgos en contextos de especies cruzadas (macaco versus humanas), de desarrollo (neonato, infantil, infantil y adulto) y clínicos (caricias perinatales) utilizando datos de PFM.
Se ha pensado que la corteza motora (M1) forma un homúnculo somatotópico continuo que se extiende por la circunvolución precentral desde el pie hasta las representaciones faciales, a pesar de la evidencia de zonas funcionales concéntricas y mapas de acciones complejas. Aquí, utilizando métodos de resonancia magnética funcional de precisión (fMRI), encontramos que el homúnculo clásico se ve interrumpido por regiones con conectividad, estructura y función distintas, alternando con áreas específicas de efectores (pie, mano y boca).
Estas regiones inter-efectoras exhiben un grosor cortical reducido y una fuerte conectividad funcional entre sí, así como con la red cingulo-opercular (CON), crítica para la acción y el control fisiológico, la excitación, los errores y el dolor. Esta interdigitación de las regiones efectoras motoras y vinculadas al control de la acción se verificó en los tres conjuntos de datos fMRI más grandes.
La IRMf de precisión de macacos y pediátricos (recién nacidos, bebés y niños) sugirió homólogos de especies cruzadas y precursores del desarrollo del sistema inter-efector. Una batería de tareas de IRMf motoras y de acción documentó somatotopías efectoras concéntricas, separadas por las regiones interefectoras vinculadas a CON.
Los interefectores carecían de especificidad de movimiento y se coactivaban durante la planificación de la acción (coordinación de manos y pies) y el movimiento axial del cuerpo (como el del abdomen o las cejas).
Estos resultados, junto con estudios previos que demuestran acciones complejas evocadas por estimulación y conectividad con órganos internos como la médula de la glándula suprarrenal, sugieren que M1 está marcado por un sistema para la planificación de acciones de todo el cuerpo, la red de acción somatocognitiva (SCAN).
En la corteza motora, dos sistemas paralelos se entrelazan, formando un patrón de integración-aislamiento: regiones específicas de efectores (pie, mano y boca) para aislar el control motor fino y el SCAN para integrar objetivos, fisiología y movimiento corporal.
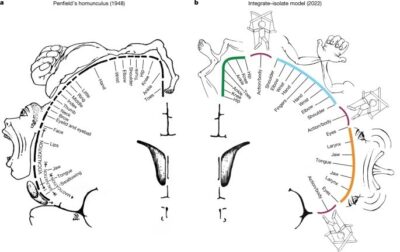
Leyenda para la Figura.
A, el homúnculo(representación de partes del cuerpo en la corteza cerebral) clásico de Penfield , que representa un mapa continuo del cuerpo en la corteza motora primaria. B, En el modelo integrado-aislado de la organización corteza motora( M1), las zonas funcionales específicas del efector (pie (verde), mano (cian) y boca (naranja)) están representadas por anillos concéntricos con partes proximales del cuerpo que rodean las distales relativamente más aislables ( dedos de manos, pies y lengua).
Las regiones inter-efectoras (granate) se asientan en los puntos de intersección de estos campos, formando parte de una red de acción somato-cognitiva para el control integrador y alostático de todo el cuerpo. Al igual que con el dibujo original de Penfield, este diagrama pretende ilustrar los principios organizacionales y no debe interpretarse en exceso como un mapa preciso.